А.Н.
Барбараш
Одесский НИИ Телевизионной Техники. Одесса, Украина
Одной из наиболее загадочных тайн, не поддающихся усилиям учёных, являлось формирование многоклеточного организма. Из единственной клетки, в ходе многочисленных делений (и в нужных случаях – уничтожений) клеток, в сочетании с непонятным науке процессом их специализации (дифференцировки), закономерно развивается очень сложная структура организма. Иногда (например, у насекомых) путь к половозрелой особи лежит через ряд других не менее сложных и законченных форм (метаморфозов). После окончания формирования организма, многие из них (например, позвоночные) длительное время (скажем, вшестеро превышающее период формирования), наперекор зависимости обмена веществ от внешней среды, сохраняют созданную структуру и восстанавливают повреждённые органы. Во все многоклеточные организмы заложены сложные и специфичные процессы создания жизнеспособного потомства. Нарушение процессов развития вызывает таинственные, мало понятные патологии – рак, трисомии, преждевременное старение детей (прогерию) и др. А после окончания генетически заданного срока жизни, организм почему-то закономерно умирает.
Все эти удивительные явления отнюдь не связаны с мистикой. Они определяются активированием в каждой клетке, в нужное время, определённых, строго отобранных генов из огромного набора, имеющегося в геноме. Упомянутые патологии, наоборот, связаны с нарушениями нормального активирования генов. Все биологические процессы, так или иначе, начинаются от активирования генов. Это хорошо известно. Но вот что не было известно – как устроен и как работает механизм активирования, каким образом в клетке выбираются и активируются именно нужные гены, составляющие часто лишь единицы или доли процентов от всего генома? Проблема управления активированием генов в многоклеточном организме, при наличии ряда бесспорных находок в этой области, оставалась для официальной науки недоступной. Поэтому биология многоклеточных организмов, составляющих более 99% известных биологических видов, и сегодня в подавляющей части оказывается лишь системой регистрации фактов, без должного теоретического объяснения. Иначе говоря, она лишена научного фундамента. Можно даже заключить, что она не вправе называться наукой.
Естественно, что такая острая проблема давно привлекала внимание многих учёных. Труднее всего оказывалось объяснить закономерное формирование сложного из простого. Подавляющее большинство предложенных концепций не объясняло этого факта. Исключением была лишь наиболее почитаемая сегодня теория диссипативных структур (ТДС), основанная на работе [Turing 1952]. Согласно ТДС, в совершенно однородной среде, в ходе особых химических реакций и процессов диффузии, может сформироваться сложное распределение концентраций сигнальных молекул – морфогенов – способное задать строение будущего организма. Важно, что предложенный теорией механизм решал трудную проблему макропликации (термин введен С.В. Македоном), т.е. формирования структуры макроуровня по информации, представленной на молекулярном уровне. Действительно, согласно ТДС, информация, заключённая в свойствах молекул, определяла параметры диффузии и химических реакций, задающие характеристики макроскопических диссипативных структур.
Математически, эта теория оказалась вполне корректной. Но она не совпала с биологическими реалиями. В организмах не был найден предсказанный теорией широкий ассортимент информационных молекул – морфогенов. Размеры животных более чем на четыре порядка превысили один миллиметр, вычисленный как предел возможных размеров их диссипативных структур. Предположению, что малый зародыш может расти уже после полного формирования, противоречило постоянство размеров диссипативных структур при постоянстве температуры. Противоречат ТДС и факты развития зародышей после образования сердца и начала кровообращения, поскольку перемещения веществ, более значительные, чем диффузия молекул, прекратили бы формирование диссипативных структур.
Необъяснимой оказалась также регенерация утраченных органов у крупных животных (например, восстановление у гигантского осьминога щупальца, откушенного кашалотом). Ведь гипотетическая диссипативная модель миллиметрового размера здесь не может помочь – организм и предполагаемая диссипативная структура отличаются масштабами в тысячи раз. Вообще, пропорциональный рост организма – один из наиболее труднообъяснимых аспектов всей проблемы развития.
С позиций ТДС непонятно, почему многоклеточными бывают только организмы с клеточными ядрами – ведь для диссипативных структур ядра не нужны. А самое заметное противоречие обнаружилось в симметрии животных – ТДС объясняет возникновение структур с осевой симметрией (морских звёзд, морских ежей и т.п.), но не объясняет формирование как раз наиболее распространённых организмов, имеющих зеркальную симметрию (позвоночных, членистоногих и др.).
Принципиальные расхождения между теорией и реальностью показали, что биологическая эволюция, действительно, могла бы пойти по пути, начертанному теорией диссипативных структур, но реально выбрала какой-то другой путь. Не случайно, через двадцать лет после работы А. Тьюринга один из наиболее авторитетных исследователей отмечал: «Именно решение проблемы наследственного осуществления информации в процессе индивидуального развития, проблем генетики развития, стало сейчас направлением главного удара … всей современной общей биологии.» [Астауров, 1972] И, вопреки последующим успехам мировой науки, ситуация до сих пор не изменилась.
После того, как стал ясен провал ТДС, в 1983 г. был найден ещё один возможный механизм макропликации. Неудача ТДС показала, что информация, управляющая развитием, по-видимому, распространяется в организме не за счёт переноса вещества, а путём переноса энергии, т.е. при помощи волнового поля. На базе этого предположения возникла концепция структурогенеза [Барбараш, 1983; Barbarash, 1992], сначала гипотетически объяснившая принципы функционирования геномов многоклеточных организмов, а затем развившаяся в подробную теорию, на этот раз точно совпавшую с реальностью. Сформировались основы новой науки – стереогенетики, разрешившей кардинальные проблемы биологии многоклеточных организмов и эукариот в целом [Барбараш, 1998].
Концепция структурогенеза (КСГ) и стереогенетика следующим образом описывают функционирование геномов эукариот.
Как предполагал Б.П. Белоусов, в протоплазме клеток эукариот периодически возникают колебательные химические реакции. В соответствии с открытием А.М. Жаботинского и А.Н. Заикина, они распространяются в виде волн [Жаботинский и др., 1988], но в данном случае, вероятно, не за счёт диффузии активных молекул, как в известных реакциях Белоусова – Жаботинского (БЖ), а с помощью иного механизма распространения. Вероятнее всего, они распространяются с помощью квантов открытого А.Г. Гурвичем ультрафиолетового излучения, которое он назвал митогенетическим. Соответственно, скорость распространения химических волн в протоплазме оценивается единицами километров в секунду, т.е. на несколько порядков более высокими значениями, чем у волн БЖ.
Предположение Б.П. Белоусова о существовании в протоплазме химических колебательных реакций косвенно подтвердилось двумя независимыми линиями экспериментов – опытами по «микродиэлектрофорезу» и «клеточному спиновому резонансу» [Pohl, 1983]. Позднее было получено подтверждение с совершенно иной стороны – сотрудник Международного института биофизики в городе Нойсе (ФРГ) Фриц Альберт Попп при исследовании митогенетического (по его терминологии – биофотонного) излучения обнаружил в нём признаки когерентности. Здесь перед нами явно новый, ранее не известный механизм возникновения когерентности. Сами по себе химические реакции не дают когерентного излучения. Свет свечи, газовой горелки или костра не когерентен. Превратить некогерентное излучение в когерентное, по-видимому, тоже невозможно – когерентность определяется синфазностью генерирования фотонов. До сих пор для получения когерентного излучения активная среда размещалась между зеркалами резонатора (интерферометра). Но в клетках организмов зеркал нет. Здесь возможен лишь один механизм синхронизации излучений атомов, способный объяснить когерентность – это синфазность молекулярных процессов и генерирования фотонов при перемещении фронта волны химических реакций. Другого объяснения когерентности УФ-излучения в данном случае найти не удаётся, поэтому сама когерентность становится весомым подтверждением существования в клетках химических волн и подтверждением положений стереогенетики.
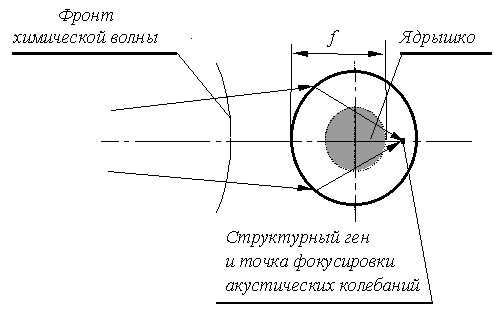
Когда передний фронт структурогенной химической волны пересекает клеточное ядро, сопутствующий ему скачок окислительно-восстановительного потенциала создаёт на оболочке ядра кольцевую зону высокой напряжённости электрического поля. Двухслойные липопротеидные мембраны выдерживают разность потенциалов до 200 мВ и более [Скулачёв, 1972; Пучкова и др., 1981]. При средней толщине каждого из слоёв 7 нм это соответствует напряжённости поля около 150 кВ/см, что близко к напряжённости пробоя лучших электротехнических изоляторов. К тому же, в отличие от технических изоляторов, липопротеидные мембраны устойчиво работают у предела электрической прочности, так как при небольших пробоях их изоляционные свойства немедленно восстанавливаются (отверстие, пробитое в молекулярной структуре, мгновенно смыкается) [Смирнов и др., 1981].
Под влиянием сильного и быстро перемещающегося электрического поля на оболочке ядра образуется «бегущая морщина» электрострикционного сжатия. Расчёты показывают, что этот процесс достаточно интенсивен – при отсутствии гидравлического сопротивления внутриядерной жидкости амплитуда «морщины» могла бы достигнуть 10% диаметра ядра. Быстрое перемещение «морщины» создаёт во внутриядерной жидкости (кариоплазме) акустические колебания. Из-за разной природы химических и акустических волн, скорости их распространения также отличаются. Оболочка ядра оказывается сферической поверхностью раздела волн с разными скоростями распространения или, с точки зрения оптики – сферической поверхностью преломления волнового поля, что придаёт ей свойства линзы.
В обычной оптике природа волн не изменяется в процессе их преломления, такую оптику можно назвать гомоволновой (одноволновой). В оптике клеточного ядра, наоборот, преломление основано на принципиальном отличии природы волн, разделяемых преломляющей поверхностью – такую оптику логично назвать гетероволновой (разноволновой).
Из-за большого разброса свойств биологических тканей следует ожидать, что интенсивность химического волнового поля в разных точках организма окажется не одинаковой – в организме неминуемо должны существовать зоны высокой и низкой интенсивности химических волн. Расчёты показывают, что в довольно широком диапазоне соотношений скоростей химических и акустических волн активные зоны химического волнового поля организма проецируются по законам оптики внутрь ядра каждой клетки в виде микроскопических зон концентрации акустических колебаний. Многочисленные факты приводят к выводу, что именно сфокусированные акустические колебания, являющиеся проекциями активных зон организма, разрыхляют хроматин в зоне расположения конкретных генов, позволяя подойти к ним ДНК-зависимым-РНК-полимеразам и осуществить транскрипцию, т.е. прочесть генетическую информацию. Так осуществляется управление активированием генов в клетках эукариот.
Стереогенетика объяснила также механизм управления дифференцировкой клеток. В геномах всех организмов содержится значительное количество обращённых (или инвертированных) повторов, иначе называемых палиндромами. Например, в организме человека палиндромы составляют более 6% общего количества ДНК [Чуанг и др., 1981]. Главная особенность палиндромов в том, что кроме обычной линейной конфигурации двойной спирали ДНК, они способны приобретать и устойчиво сохранять иную – крестообразную конфигурацию. Палиндромы выполняют в геноме разные функции, например, небольшие палиндромы встроены в терминаторные участки генов. Но есть группа крупных палиндромов, управляющих процессами дифференцировки клеток. В оплодотворённой яйцеклетке эти палиндромы находятся в исходном линейном состоянии. Когда на них воздействуют сфокусированные акустические колебания и соответствующий участок хроматина перейдёт в разрыхлённое состояние, к ним получают доступ расплетающие и релаксирующие ферменты (хеликаза и ДНК-гираза). Под их воздействием двойная спираль ДНК расплетается на две нити и появляется возможность случайного выбора – процессы самосборки могут снова собрать нити в линейную двойную спираль, но могут привести и к крестообразной конфигурации. Вероятность сборки в форме креста не одинакова для разных палиндромов и тем выше, чем короче внутренний спейсер палиндрома.
Вероятностное изменение конфигурации палиндрома после воздействия волнового поля и расплетения ДНК не изменяет пространственного расположения генов в данной клетке, потому что ДНК эукариот во множестве точек прикреплена к ядерному белковому матриксу и к оболочке ядра. Соответственно, в данной клетке не изменяются и условия активирования генов. Но палиндромы способны сохранять свою конфигурацию при репликации и переходить в дочерние клетки, а так как самосборка хроматина очень чувствительна к конфигурации ДНК, то каждый новый крест ведёт к новому пространственному расположению генов в ядре дочерней клетки. При этом, по сравнению с родительской клеткой, одни гены покидают активные зоны акустического волнового поля ядра и переходят в неактивное состояние, а другие гены, напротив, входят в активные зоны и становятся доступными для активирования волновым полем. Так происходит устойчивое изменение свойств клеток в процессе их дифференцировки.
Оптический механизм активирования генов обладает яркой спецификой, рельефно проявляющейся в реальных организмах. Его свойства невозможно спутать со следствиями ТДС или любой другой концепции. Так, в отличие от прокариот, для чёткой работы оптического механизма гены и палиндромы должны быть стабильно зафиксированы в пространстве ядра, чего не требует ТДС, но что, действительно, обеспечивается многоточечным прикреплением хромосом к белковому матриксу ядра и к ядерной оболочке.
Или другой пример. Фокусное расстояние клеточного ядра f составляет:
f
= nR / (n – 1),
(1)
где R – радиус ядра, n – гетероволновый коэффициент преломления, т.е. отношение скоростей химических и акустических волн (для многоклеточных организмов n ≈ 2.6–2.8, а для одноклеточных эукариот n ≈ 3.5). Из формулы вытекает, что в центральной части ядра есть так называемая безадресная сферическая область, в которую не могут проецироваться никакие внеядерные точки. Радиус r безадресной области равен:
r
= R / (n – 1),
(2)
В этой области не могут располагаться структурные гены, активируемые проекциями активных зон организма. И, действительно, в центральной части ядра располагается ядрышко (ожидаемого размера), содержащее только гены рибосом, которые не относятся к группе структурных!
Оптические расчёты показывают также, что от оболочки ядра к границам безадресной области нелинейно растёт вероятность проекции активных зон организма в единицу объёма хроматина, достигая вблизи границ безадресной области очень высоких значений. Это совпадает с фактическим распределением активного хроматина (эухроматина) в ядре – он сильнее всего сконцентрирован вокруг ядрышка (что непонятно для ТДС), а к ядерной оболочке всё более уступает место неактивному гетерохроматину. Характерно, что и тельца Барра (лишние Х-хромосомы женщин) укрываются от активирования именно вблизи ядерных оболочек, где вероятность активирования минимальна.
Оптический подход объяснил
также удивительный факт (снова-таки,
непонятный для ТДС) концентрации наиболее
активных генов – генов рибосом (дающих
около 80% всех РНК) в центральной части ядра,
что резко усложнило транспорт из
цитоплазмы исходных веществ и перемещение
РНК в цитоплазму. Причина же оказалась в том,
что все акустические волны, активирующие
структурные гены, прежде, чем
сфокусироваться на гене, проходят в
расфокусированном состоянии сквозь
ядрышко, как это показано на рис. 1.
Рис. 1. Схема активирования структурного гена
Это приводит к пропорциональности между средней интенсивностью волнового поля в области ядрышка и суммарной потребностью клетки в синтезе белков, то есть потребностью клетки в рибосомах. Природа использовала такой удобный случай для управления активностью генов рибосом, разместив их в этой области. А поскольку расфокусированные волны менее интенсивны, хроматин ядрышка приспособился к разрыхлению слабыми колебаниями – щелочные белки-гистоны заменены в хроматине ядрышка более кислыми белками, слабее связывающимися с ДНК (но зато лучше удерживающими основные красители, что и выделило ядрышко как морфологическое образование).
Потребность в волновом управлении работой генома возникает к моменту начала транскрипции. Поэтому на первых этапах развития зародыша, например, у амфибий, пока не начат синтез собственных РНК (используются ранее заготовленные материнские РНК), управление с помощью волнового поля отсутствует. Это означает отсутствие у клеток информации об их расположении в структуре зародыша и приводит к одинаковому поведению всех клеток, в том числе, к синхронности делений.
Переход от синхронного к асинхронному делению говорит о первом всплеске структурогенного волнового поля, о том, что оно взяло «бразды правления» на себя. Например, у лягушки Xenoрus laevis первые 12 циклов деления проходят синхронно, а затем внезапно, казалось бы – без всяких причин, синхронность нарушается, и зародыш переходит к асинхронному делению, характерному для всей последующей жизни. Это и есть момент «пробуждения» волнового поля. Об этом говорит, в частности, поведение генов рибосом. На этапе синхронного деления клеток зародыша лягушки тысячи рибосомальных генов распределены у оболочек ядер, что наиболее благоприятно для транспорта исходных веществ из цитоплазмы и для противоположного перемещения рРНК. «Переход к асинхронному делению сопровождается рядом цитологических перестроек, из которых самая наглядная – появление нормального ядрышка в центре ядра каждой клетки» [Белоусов, 1980]. Формирование ядрышка – это и есть переход генов рРНК под управление волновым полем.
Для оптического управления работой генов характерно, что основные процессы воздействия волнового поля на гены приурочены к неподвижности организма, к одной из фаз сна. Такая же связь активирования генов с относительной неподвижностью видна и на клеточном уровне – в кувыркающихся клетках жидкостей внутренней среды животных структурные гены выключены, вплоть до того, что у млекопитающих более 99% клеток кровотока (все красные клетки крови) вовсе лишены ядер, что совершенно непонятно с позиций ТДС.
Хотя много десятилетий идёт речь об изучении «морфогенеза», т.е. генезиса форм тел живых организмов, такой подход лишён смысла. Применительно к животным вообще нельзя говорить об определённых формах; у них нет стабильности форм; никто не скажет, например, какая поза человека является его главной, видовой позой. Поэтому искать механизмы наследования форм организмов методологически ошибочно. Нет оснований считать, будто организм наследует от предков определённую форму. Наследуется структура (анатомия) тела, а не его форма.
Как бы это ни показалось унизительным для человека, наш организм похож не столько на мраморную статую Аполлона, сколько на невзрачную сетку-«авоську». Попробуйте изменить позу мраморной статуи – и вы сломаете её. Ничего подобного не произойдёт с «авоськой». Она может стать шарообразной, если вместила арбуз, может жгутом висеть на крючке или комочком лежать в кармане. При этом форма «авоськи» меняется в широких пределах. Но пока сетка цела, она сохраняет неизменную схему расположения узелков сети, неизменные длины нитей между узелками, и можно говорить о её неизменной структуре. Человеческий организм (и любой другой многоклеточный организм) тоже характеризуется определённой структурой. И именно структура организма кодируется генетическим аппаратом. Если точнее, то координатами генов в ядре и их взаимодействием с волновым полем задаются векторы (процессы) формирования структуры организма.
Чтобы организм формировался генетической системой именно как структура, он должен обладать определёнными свойствами, как среда распространения химических волн. Нужно, чтобы параметры волнового поля организма определялись не столько проходимым волной расстоянием, сколько особенностями структуры, по которой волна распространяется. Реальный характер связей между соседними клетками создаёт именно такие условия. Переход волны химических реакций от клетки к клетке происходит через множество коннексонов – трубочек молекулярного размера, пронизывающих стенки клеток. Суммарная площадь поперечного сечения каналов коннексонов, соединяющих клетки, на несколько порядков меньше площади поперечного сечения самой клетки. Поэтому сопротивление прохождению химической волны определяется, почти исключительно, неизменяемым числом пройденных переходов от клетки к клетке и стабильным количеством коннексонов на каждом переходе, а не размерами клеток, деформируемых при изменениях позы. Потому-то изменения позы мало влияют на параметры структурогенного волнового поля. Потому-то структурогенное волновое поле описывает (и прочитывает из генома) структуру, а не форму организма.
Однако нельзя считать, что информационный результат прохождения химических волн по биологическим тканям совершенно не зависит от расстояния. Если бы геометрическая длина пути никак не отражалась на информационных характеристиках химических волн, то движения животного нисколько не мешали бы процессам развития, и тогда животным не нужен был бы сон, как физиологическое состояние (как он не нужен растениям). Но необходимость в периодах неподвижности, необходимость периодического сна для точного активирования генов сохранилась.
Рост организма слегка смещает проекции активных зон в ядрах клеток, что может прервать активирование нужных генов. Как противодействие такому явлению, структурные гены обычно присутствуют не поодиночке, а цепочками (кластерами) однотипных генов, последовательно включаемых и выключаемых по мере перемещения проекции активной зоны (т.е. по ходу жизни организма). Поскольку активирование генов волновым полем безразлично к ориентации гена, то гены кластеров, при строгой очерёдности расположения, ориентированы произвольно (в отличие от генов оперонов прокариот). Возможность нормального управления активированием генов и дифференцировкой клеток в процессе роста организма, как выяснилось, обеспечивается тремя факторами:
· записью в геноме не статичных форм организма, а векторов развития;
· существованием кластеров генов;
· особенностями оптической проекции удалённых зон организма на геномы клеток, при которой значительные вариации расстояний до активных зон не нарушают точности активирования генов (как фотоаппарат резко изображает все объекты, удалённые более, чем на 20 м).
Чтобы в пространстве ядра точно совместить гены с проекциями активных зон волнового поля организма, нужна неинформативная ДНК в виде вставок между генами. Реальные геномы эукариот, действительно, имеют такую, казалось бы, ненужную фракцию. Кроме того, в разных зонах организма одинаковые гены (например, мышц, артерий и т.п.) должны функционировать при разной картине волнового поля, что требует повторения их во многих точках ядерного пространства. Это объяснило непонятное для ТДС многократное повторение в геномах эукариот одинаковых структурных генов.
Стереогенетика объяснила также широкий круг других непонятных ранее явлений. Например, она указала, что нарушения пространственного согласования генома с проекциями активных зон волнового поля (из-за трисомий, деформаций ядра, внедрения вирусов, нарушения пространственной структуры ДНК и т.п.) ведут к тяжёлым последствиям для организма. Действительно, все подобные факторы с высокой вероятностью вызывают смерть зародыша либо тяжёлые патологии, часто – канцерогенез. Очень показательно закономерное возникновение злокачественной опухоли при имплантации зеркальной пластины в брюшную полость крысы – отражение химических волн от зеркала вызывает образование аномальных «стоячих волн». Такой механизм патологии подтверждается тем, что первая злокачественно переродившаяся клетка, дающая начало всему клону клеток опухоли, никогда не появляется в контакте с вызвавшей её зеркальной пластиной, а всегда – на некотором расстоянии.
Исследователи химических волн полагают, что такое явление невозможно – мол, химические волны не способны отражаться и интерферировать. Но подобное возражение справедливо лишь для химических волн с малыми молекулами, имеющими близкий к единице стерический фактор реакции (например, для волн БЖ). При участии в реакции крупных органических молекул стерический фактор падает до долей процентов, отчего становится возможным и отражение, и интерференция волн.
По ходу эволюции, центр тяжести генетических изменений перемещался с молекулярного строения генов на изменения в длинах некодирующих участков ДНК, на характер расположения генов в пространстве ядра. Обнаружилось соответствие между темпами изменений структур генома и организма в целом. У высоко развитых таксонов (например, у приматов, включая людей), большинство различий геномов касаются уже не генов, а их пространственного расположения. Этот факт, непонятный для теории диссипативных структур, естественно вытекает из положений стереогенетики.
Перечень можно продолжать и продолжать. Положения стереогенетики совпали с биологическими реалиями всюду, где только их можно сопоставить, вплоть до тонких нюансов. Эта удивительная наука выявила, в частности, анатомическую особенность млекопитающих, затруднившую регенерацию утраченных конечностей. На этой основе намечен способ восстановления конечностей, обещающий излечение многих инвалидов. Стереогенетика – непростая наука, о которой одна статья даёт лишь поверхностное представление. Подробнее о ней можно прочесть в книге [Барбараш, 1998] или (в более поздней редакции) в Интернете, в работах «Основы стереогенетики», «Код. Жизнь. Вселенная. (Теории и гипотезы)» и др. на персональном сайте автора: http://barbarashan.narod.ru/.
Выводы стереогенетики во многом неожиданны и парадоксальны. С позиций стереогенетики, организм человека содержит миллиарды оптических систем, по разрешающей способности и масштабному коэффициенту эквивалентных электронным микроскопам! Клетка определяет своё положение в организме по окружающим активным зонам волнового поля, как раньше моряки определяли координаты корабля по звёздам. Удивительно! Но сомнений в этом сегодня уже нет. Стереогенетика стала теоретическим фундаментом, поднявшись на который биология многоклеточных организмов превратится из системы регистрации фактов в подлинную науку.
Почему этого не произошло до сих пор? Почему стереогенетика всё ещё не стала достоянием мирового сообщества? Причина ясна. Как справедливо указывал Андрей Везалий, «природа не разделена на факультеты, подобно нашим университетам». В стереогенетике, изучающей одну из сложнейших загадок природы, особенно тесно сплелись биологический, оптический, кибернетический, волновой, химический, медицинский и другие аспекты. Поэтому в судьбе стереогенетики особенно ярко проявилось фундаментальное противоречие познания – наука беспредельна, а объём человеческого мозга ограничен. Ведь и университеты разделились на факультеты не случайно! Всё более узкая специализация – это способ борьбы среднего землянина с необъятностью науки.
Как раз из-за соединения в стереогенетике многих аспектов науки, ни один из узких специалистов (особенно, биологов) её не понял. Узость кругозора рецензентов помешала публикации журнальных статей. Опыт показал, что понять и оценить стереогенетику способен лишь коллектив разнопрофильных учёных. Но немногие существующие комплексные научные коллективы заняты собственными работами. Никому не хочется отвлекаться на что-то чужое и сложное, будь то хоть важнейшая из проблем. Потому-то стереогенетика оказалась наукой XXI, а не XX века. Можно лишь гадать, каких потерь удалось бы избежать, скольким инвалидам (скажем, африканским детям, пострадавшим от противопехотных мин) можно было бы вернуть радости жизни, если бы непроходимое болото узкой специализации учёных не отгородило эти работы от широкой научной общественности.
Л И Т Е Р А Т У Р А
Астауров Б.Л. Генетика и
проблемы индивидуального развития //
Онтогенез. Т. 3.
1972, N 6. С.
547–549.
Барбараш А.Н. Гипотеза о механизме активирования генов эукариот в связи с канцерогенезом. Киев, 1983. 23 с. Деп. в УкрНИИНТИ, 22.11.83, N 1313 Ук-Д83.
Барбараш А.Н. Волновые процессы в живом: основы стереогенетики и физиологии мышления. Одесса: ОМ, ПОЛИС. 1998. 352 с.
Белоусов Л.В. Введение в общую эмбриологию. М.: Изд-во МГУ. 1980. 216 с.
Жаботинский А.М., Отмер Х., Филд Р. и др. Колебания и бегущие волны в химических системах. М.: Мир. 1988. 720 с.
Пучкова Т.В., Путвинский А.В., Владимиров Ю.А. Электрический пробой бислойных фосфолипидных мембран диффузионным калиевым потенциалом. // Биофизика. 1981. Т. 26, вып. 2. С. 378–379.
Скулачёв В.П. Трансформация энергии в биомембранах. М.: Наука, 1972.
Смирнов А.А., Путвинский А.В., Рощупкин Д.И., Владимиров Ю.А. Необратимое повреждение липидных бислойных мембран в состоянии электрического пробоя. // Биофизика, 1981. Т. 26, вып. 1. С. 140–141.
Чуанг Ц.Р., Дотт П.Дж., Тьен
Куо М., Сандерс Г.Ф. Обращенные
повторяющиеся последовательности в ДНК
человека. // Молекулярные основы генетич.
процессов. Труды XIV Междунар. генетич.
конгресса. Москва 21-30 авг. 1978. М.: Наука. 1981.
С. 362–369.
Barbarash
A.N. The Concept
of Structuregenesis // Acta Biotheoretica. 1992. V. 40 (1). P. 89–90.
Pohl
H. A natural
oscillating fields of cells // Coherent Excitad. Biol. Syst. Berlin a.e. 1983, p.
199–210.
Turing A.M. The chemical basis of the morphogenesis // Proc. Roy. Soc. B. 1952. V. 273. P. 37–71.
Стереогенетика раскрыла механизм активирования генов эукариот. В протоплазме возникают волны химических колебательных реакций. Передний фронт волны несёт скачок окислительно-восстановительного потенциала, и вызывает электрострикционную деформацию оболочки клеточного ядра, порождая акустические колебания внутриядерной жидкости. Различие скоростей распространения химических и акустических волн вызывает преломление волнового поля на ядерной оболочке, отчего она приобретает свойства линзы и проецирует активные зоны химического волнового поля организма на хроматин ядра в виде микроскопических зон акустических колебаний. Они и разрыхляют хроматин в зонах, определяющих активирование генов и дифференцировку клеток.
The stereogenetics has
discovered a mechanism of eucaryotic genes activation. The waves of chemical
oscillatory reactions arise in protoplasm. The forward front of a wave carries a
leap of a redox potential and causes an electrostrictional deformation of
cell nuclear membrane, thus generating the acoustic oscillations of
intranuclear liquid. The diffference in velocity of spread of chemical and
acoustic waves produces a refraction of wave field on the nuclear
membrane, so that the membrane acquires the properties of a lens and projects
the active zones of a chemical wave field of the organism onto the
nuclear chromatin as the microscopic zones of acoustic oscillations focus. They,
in their turn, loosen the chromatin in the zones which determine
genes activation and cells differentiation.